Starling equation
The Starling equation is an equation that illustrates the role of hydrostatic and oncotic forces (the so-called Starling forces) in the movement of fluid across capillary membranes.
Capillary fluid movement may occur as a result of three processes:
- diffusion
- filtration
- pinocytosis
Starling's equation only refers to fluid movement across the capillary membrane that occurs as a result of filtration. In the glomerular capillaries, there is a net fluid filtration of 125 ml/min (about 180 litres/day). In the rest of the body's capillaries, there is a total net transcapillary fluid movement of 20 ml/min (about 28.8 litres/day) as a result of filtration. This is several orders of magnitude lower than the total diffusional water flux at the capillary membrane, as that is about 80,000 litres/day.
The Starling equation was formulated in 1896 by the British physiologist Ernest Starling, also known for the Frank–Starling law of the heart.
Contents |
The equation
The Starling equation reads as follows:
![\ J_v = K_f ( [P_c - P_i] - \sigma[\pi_c - \pi_i] )](/2012-wikipedia_en_all_nopic_01_2012/I/5768210d96d6bfb8a9742b3861756b66.png)
where:
![( [P_c - P_i] - \sigma[\pi_c - \pi_i] )](/2012-wikipedia_en_all_nopic_01_2012/I/c944a55e63e74323ad5ee4c7073042b6.png) is the net driving force,
is the net driving force,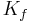 is the proportionality constant, and
is the proportionality constant, and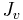 is the net fluid movement between compartments.
is the net fluid movement between compartments.
By convention, outward force is defined as positive, and inward force is defined as negative. The solution to the equation is known as the net filtration or net fluid movement (Jv). If positive, fluid will tend to leave the capillary (filtration). If negative, fluid will tend to enter the capillary (absorption). This equation has a number of important physiologic implications, especially when pathologic processes grossly alter one or more of the variables. Note that previously it was believed that at steady state the arterial capillaries filter fluid and the venous capillaries reabsorb it, as shown by the diagram. Though many physiology textbooks still use this misconception, modern evidence shows that in most cases venular blood pressure exceeds the opposing pressure, thus maintaining a positive outward force. This indicates that capillaries are normally in a state of filtration along their entire length. [2]
The variables
According to Starling's equation, the movement of fluid depends on six variables:
- Capillary hydrostatic pressure ( Pc )
- Interstitial hydrostatic pressure ( Pi )
- Capillary oncotic pressure ( πc )
- Interstitial oncotic pressure ( πi )
- Filtration coefficient ( Kf )
- Reflection coefficient ( σ )
Pressures are often measured in millimetres of mercury (mmHg), and the filtration coefficient in millilitres per minute per millimetre of mercury (ml·min-1·mmHg-1).
In essence the equation says that the net filtration (Jv) is proportional to the net driving force. The first four variables in the list above are the forces that contribute to the net driving force.
Filtration coefficient
The filtration coefficient is the constant of proportionality. A high value indicates a highly water permeable capillary. A low value indicates a low capillary permeability.
The filtration coefficient is the product of two components:
- capillary surface area
- capillary hydraulic conductance
Reflection coefficient
The reflection coefficient is often thought of as a correction factor. The idea is that the difference in oncotic pressures contributes to the net driving force because most capillaries in the body are fairly impermeable to the large molecular weight proteins. (The term ultrafiltration is usually used to refer to this situation where the large molecules are retained by a semipermeable membrane but water and low molecular weight solutes can pass through the membrane).
Many body capillaries do have a small permeability to proteins (such as albumins). This small protein leakage has two important effects:
- the interstitial fluid oncotic pressure is higher than it would otherwise be in that tissue
- not all of the protein present is effective in retaining water so the effective capillary oncotic pressure is lower than the measured capillary oncotic pressure.
Both these effects decrease the contribution of the oncotic pressure gradient to the net driving force. The reflection coefficient (σ) is used to correct the magnitude of the measured gradient to 'correct for' for the ineffectiveness of some of the oncotic pressure gradient. It can have a value from 0 up to 1.
- Glomerular capillaries have a reflection coefficient close to 1 as normally no protein crosses into the glomerular filtrate.
- In contrast, hepatic sinusoids have a low reflection coefficient as they are quite permeable to protein. This is advantageous because albumin is produced in hepatocytes and can relatively freely pass from these cells into the blood in the sinusoids. The predominant pathway for albumin and other proteins to enter the circulation is via the lymph.
Approximated values
Following are approximated values for the variables in the equation for both arterioles and venules:
| Location | Pc (mmHg) [3] | Pi (mmHg) [3] | σπc (mmHg) [3] | σπi (mmHg) [3] |
|---|---|---|---|---|
| arteriolar end of capillary | +35 | -2 | +28 | +0.1 |
| venular end of capillary | +15 | -2 | +28 | +3 |
Some albumin escapes from the capillaries and enters the interstitial fluid where it would produce a flow of water equivalent to that produced by a hydrostatic pressure of +3 mmHg. Thus, the difference in protein concentration would produce a flow of fluid into the vessel at the venous end equivalent to 28 − 3 = 25 mmHg of hydrostatic pressure. The total oncotic pressure present at the venous end could be considered as +25 mmHg.
In the beginning (arteriolar end) of a capillary, there is a net driving force () outwards from the capillary of +9 mmHg. In the end (venular end), on the other hand, there is a net driving force of -8 mmHg.
Assumed that the net driving force declines linearly, then there is a mean net driving force outwards from the capillary as a whole, which also results in that more fluid exits a capillary than re-enters it. The lymphatic system drains this excess.
Clinical usefulness
The equation is very useful for explaining physiological phenomena happening at the capillary (e.g. the formation of edemas), but has very limited clinical usefulness. Mostly this reflects the impossibility of easily measuring all six variables together in actual patients.
See also
References
- ^ West, John (2012). Respiratory Physiology : the essentials - 9th edition. Baltimore: Lippincott Williams & Wilkins. pp. 177. ISBN 978-1-60913-640-6.
- ^ Levick J.R., Introduction to Cardiovascular Physiology. Oxford Press, 2003, p. 179-180.
- ^ a b c d Walter F., PhD. Boron. Medical Physiology: A Cellular And Molecular Approaoch. Elsevier/Saunders. ISBN 1-4160-2328-3.